EMERGENTE EVOLUTIE*
Zonder taal zijn verhalen onbestaanbaar
Mijn diepgelovige buren zijn er niet langer van overtuigd dat evolutie iets is voor goddelozen: amoreel en willekeurig. Ook in hún kring van godvrezende kerkgangers gaan remonstrantse stemmen op dat God en evolutie elkaar niet hoeven uit te sluiten. Wetenschap (evolutie) is een geschenk van God, die hemel en aarde schiep in zeven dagen. Nou ja, dat laatste hoeven ze ook niet letterlijk te nemen. God schiep de oerknal, waar alles uit is voortgekomen en waarmee Hij schuilgaat achter het hele bestaan. Zijn roep vanuit de hemel leidt hen vanzelf naar het Goede want Hij is ook heel dichtbij. In henzelf geeft Hij hen hoop en vertrouwen.
De verhalen in de Bijbel wekken de indruk van een geschiedenis. Een begin (de schepping), middenmoot (stamvaders; leven van Jezus) en een einde (profetie over de eindtijd). Die historische lijn is echter niet origineel. Het begin is later geschreven dan de geschiedenis van de stamvaders. Het verhaal van een scheppende God diende als steun in onzekere tijden.
Persoonlijk heb ik die verhalen niet zo nodig om te begrijpen hoe de wereld in elkaar steekt. Een wetenschappelijke verklaring voldoet. Emoties als hoop, vertrouwen en liefde (en scheidend vermogen tussen goed en kwaad) komen van binnenuit en worden geïnitieerd door hormonen*
Bij evolutie denken de meeste mensen aan het ontstaan van nieuwe planten en dieren uit eerdere levende wezens. Soms is er zelfs een notie van gemeenschappelijk voorouderschap en natuurlijke selectie. Niet iedereen is het eens met de ideeën van Charles Darwin maar zijn naam kennen ze allemaal.
Minder vaak wordt evolutie in verband gebracht met de ontwikkeling van niet-levende materie. Zoals de evolutie van planeten en sterren, de kosmologische ontwikkelingen, het ontstaan van het heelal. Niet iedereen is bekend met de ideeën van Stephen Hawking maar zijn naam kennen ze allemaal.
Elke nieuwe fase van evolutie bouwt voort op een enorme keten van toevalligheden die verankerd zijn geraakt. Zo bepalen lagere niveaus van evolutie de contouren van de omgeving waarin hogere niveaus zich kunnen ontwikkelen. Dit laat zoveel ruimte voor verrassende wendingen dat buitengewoon onwaarschijnlijke paden vaak werkelijkheid worden en determinisme faalt. De toevallige uitkomsten bij ontelbare vertakkingen brengen een emergent element binnen in de evolutie. Ze voegen een grote hoeveelheid informatie en structuur toe die simpelweg niet in de lagere niveaus te vinden zijn, van waaruit wetten op een hoger niveau tevoorschijn kunnen komen - en dat doen ze ook vaak. Zo gelooft bijvoorbeeld geen enkele serieuze wetenschapper vandaag dat er in de biologie speciale 'levenskrachten' bestaan die geen fysio-chemische grondslag hebben, maar de fysische wetmatigheden alleen bepalen geenszins wat de wetten van de biologie op aarde zullen zijn.*
Om de enorme verscheidenheid aan soorten te verklaren definieerde Darwin in 1859 het mechanisme van de natuurlijke selectie in relatie tot seksuele voortplanting. In 1937 formuleerde Dobzhanski de ‘modern synthesis’ met natuurlijke selectie als kern van de genetische evolutie. Decennia lang werd aangenomen dat natuurlijke selectie werkte op het niveau van soorten en zelfs van ecosystemen, totdat duidelijk werd dat de kracht ervan veel groter was op het niveau van het individuele organisme en volgens Dawkins zelfs nog sterker op het niveau van genen (1976). Het mechanisme van natuurlijke selectie bleek op genetisch niveau krachtig genoeg om organische evolutie te verklaren, zonder doel of toekomstige attractor. Wederkerig altruïsme en speltheorie suggereerden het bevorderen van de samenwerking tussen totaal verschillende soorten, wat leidde tot een betere verklaring van de evolutie van altruïsme. Met de introductie van de ‘major transitions’ in de evolutie ontstond een heropleving van de multilevel (groeps) selectie met veelbelovende nieuwe inzichten.*
Tweehonderd jaar geleden verzamelde de elfjarige Charles Darwin kevertjes in de tuin van het ouderlijk huis en verwonderde zich over de rijkdom van Gods Schepping. Op zoek naar inzicht stuitte hij in zijn vaders boekenkast op James Huttons Theory of the Earth (1795) en maakte kennis met het uniformitarianisme (het heden is de sleutel tot het verleden). Daar is wellicht de kiem gelegd om met andere ogen naar de wereld te kijken. Met de publicatie van het principe van natuurlijke selectie dat achter zijn verwondering schuilging heeft hij overigens lang getalmd, toen het verscheen in 1859 was hij vijftig jaar oud. Sindsdien bestaat er een intellectuele tweespalt tussen het geloof in doelgerichte schepping en de claim op toevallige ontwikkeling. De eerste stroming vertegenwoordigt alle wereldreligies en (pseudo)wetenschappelijke opvattingen zoals creationisme (incl. intelligent design) en theïstisch evolutionisme. De andere, veel kleinere, groep omvat de agnostische wetenschap. De vraag is of de onmiskenbare toename van complexiteit het gevolg is van toeval of van doelgerichtheid.
Meer dan drie miljard jaar organische evolutie toont onweerlegbaar de cruciale rol van gecoördineerde actie en samenwerking. De ‘major transitions’ in de evolutie vertegenwoordigen complexiteitssprongen die hun oorsprong vinden in samenklontering. Het is een plausibel idee dat het geringe aantal innovatieve gebeurtenissen in de organische evolutie het gevolg is van groepsselectie in plaats van individuele selectie. Het gaat immers om de overerving van emergente kenmerken door differentiële groepsproductiviteit. Aan het begin van elke transitie ontstond er als het ware een 'maagdelijke' dimensie voor de emergente indringers die zich zonder concurrentie snel konden ontwikkelen totdat natuurlijke selectie hen dwong tot adaptatie. Volgens Kauffman is natuurlijke selectie misschien niet eens de belangrijkste bron van biologische organisatie, maar meer een soort fijnregeling binnen een zelforganiserende natuurlijke wereld. Groepsselectie lijkt elke keer dat er een transitie ophanden is de individuele selectie uit evenwicht te brengen.
In de meeste biologiemethodes van het voortgezet onderwijs wordt het evolutiemodel ingedikt tot natuurlijke selectie en populatiegenetica in het dierenrijk. De andere meercellige organismen (planten en schimmels) spelen een ondergeschikte rol, evenals overigens de eencelligen. Een overzicht van de ontwikkeling van het leven op aarde ontbreekt niettemin zelden. Om de geologische tijdschaal wat aansprekender te maken wordt deze wel eens gepresenteerd als een klok: de mens ontstond om 1 seconde voor klokslag 12 uur, één minuut eerder verdwenen de dino’s en kwamen de zoogdieren op, de meercelligen (planten, dieren en schimmels) kwamen om 10 vóór 12 tot ontwikkeling, voorafgegaan door de eencellige eukaryoten die zich voor het eerst lieten zien rond half 12. Het eerste half uur was volledig gevuld met prokaryoten ((arche)bacteriën). Misschien dat deze uit levenloze materie zijn ontstaan (op aarde of elders) maar dát was in elk geval vóór elven. Opvallend is dat het hoofdstuk Evolutie focust op die laatste minuut. Een kritische leerling zal zich afvragen of natuurlijke selectie en populatiegenetica over de hele tijdschaal geldt. Vooral op de in het oog springende momenten waarop er iets wezenlijks verandert. Wat is daar aan de hand? Telkens als die vraag rijst moet ik denken aan de kritische leerling die zich had verwonderd over de vorm van Afrika en Amerika waardoor de continenten op aansluitende puzzelstukjes lijken (in An Inconvenient Truth van Al Gore). Ook de ontwikkeling van het leven op aarde vertoont namelijk opmerkelijke scheilijnen. Uitgezonderd de endosymbiontentheorie (ontstaan van eukaryoten uit symbiotische prokaryoten) wordt in geen enkele methode aandacht besteed aan die evolutionaire reuzensprongen. In 1995 publiceerden Szathmary en Maynard Smith in Nature voor het eerst een lijstje van major transitions in evolution. Zij onderscheidden een achttal overgangen: ‘symbiose’ tussen replicerende en membraan vormende moleculen; clustering van replicerende moleculen tot chromosomen; ontstaan van de genetische code; endosymbiose; overgang van aseksuele klonen naar seksuele populaties; overgang van protist naar meercelligen (differentiatie); overgang van solitair naar kolonie; ontstaan van de talige humane samenleving. Aan de hand van uiteenlopende criteria kon hun lijstje worden veranderd of uitgebreid (met bv. de kolonisatie land door korstmos; ontstaan van het immuunsysteem). Eén ding hebben die lijstjes gemeen: er is sprake van een toenemende complexiteit.
Evolutionaire transities hebben een aantal gemeenschappelijke kenmerken. Elke transitie verschaft een reeks volledig nieuwe attributen. Daarbij is de entiteit of het systeem zowel in complexiteit als in omvang met minstens een factor 10 toegenomen. De transitie kan worden geïnterpreteerd als een stap naar een volgend organisatieniveau. Om de transitie mogelijk te maken vormen de betrokken componenten een band met voldoende overlevingskans om het volgende niveau te betreden. De componenten zijn in principe niet identiek, ze zijn zelfs heel verschillend (als er veel componenten betrokken zijn kunnen ze in groepen voorkomen) en hun oorspronkelijke 'vrijheid' wordt omgevormd tot een onderlinge alliantie. Kenmerkend voor transities is dat ze iets nieuws toevoegen en niet iets vervangen dat al bestaat (de term ‘revolutie’ voor dergelijke veranderingen is dus pertinent onjuist). In feite interageren de nieuwe systemen met de reeds bestaande organisatieniveaus, wat nog eens een extra dimensie van complexiteit met zich meebrengt.
Een systeem wordt emergent genoemd als de componenten innovatieve kwaliteiten hebben die het systeem in staat stellen 'een nieuwe dimensie te veroveren'. Synergie speelt een sleutelrol bij de ontwikkeling van complexe systemen, vooral met betrekking tot hun samenwerking. In de (organische) evolutie kan het worden opgevat als een functioneel principe met betrekking tot de overlevingskans van emergente systemen. In het kader van niet-lineaire dynamische systeemmodellen beschouwt Kauffman synergie (emergentie) als de uitkomst van een zelforganiserend systeem met binaire subsystemen, balancerend tussen een chaotische en een sterk geordende toestand.
Er is sprake van synergie bij de vorming van elementaire deeltjes, atomen en moleculen, bij het tot stand komen van chemische verbindingen, bij het ontstaan van leven, bij de endosymbiotische vorming van eukaryote cellen, bij de ontwikkeling van meercellige organismen, bij de verovering van het land door terrestrische symbionten, bij socialisatie door individuele organismen en bij de wereldwijde communicatie via elektronische netwerken. In managementadviesbureaus wordt synergie vaak gebruikt om de voordelen van samenwerking aan te geven. Vaak wordt op deze voordelen gewezen om bedrijfsfusies, meestal ten koste van het personeel, te verdedigen. Als het om verschillende biologische soorten gaat gebruikt men liever de term symbiose. Synergie wordt ook gebruikt als een overkoepelende term voor allerlei soorten onherleidbare coöperatieve effecten. Dit zou gemakkelijk tot misverstanden kunnen leiden, omdat creationisten geloven dat onherleidbare complexiteit het bestaan van Intelligent Design bevestigt. Onherleidbaarheid is echter een illusie. De onmogelijkheid om complexe structuren te vereenvoudigen wordt veroorzaakt door dogmatisch denken en gebrek aan creativiteit. Niet-lineaire oscillaties lijken alleen maar onvoorspelbaar. Zodra je doorhebt wat er aan de hand is zijn de oorzaken, weliswaar achteraf, eenvoudig te begrijpen. Zolang een causale verklaring zoals ‘natuurlijke selectie’ de zoeker inzicht verschaft, is er geen logische noodzaak om op zoek te gaan naar een goddelijke ontwerper of Bijbelse verklaring.
Het blinde toeval, dat op het onderliggende genetische mechanisme van de natuurlijke selectie van toepassing is, geldt waarschijnlijk niet alleen voor dieren maar ook voor eencellige protisten en bacteriën. Voor de transities naar een hoger plan, het opgeven van de eigen identiteit ten behoeve van een innige samenwerking, is echter meer nodig. Dat schijnbaar doelgerichte streven naar grotere complexiteit prikkelt sommigen tot de religieuze aanname van een anti-causale, uit de toekomst afkomstige aantrekkingskracht die het evolutieproces voorttrekt. Terecht noemt Klapwijk (2009) de transitie een voorbeeld van emergentie, een nieuwe zijnsvorm die niet simpel valt te herleiden tot de eraan voorgaande componenten. Maar dat betekent nog niet dat zo’n emergentie geen causale ontstaansgeschiedenis kent (zie Van Belle & van der Veken, 2008). Door de structuren tijdens hun transitie te zien als complexe systemen met zelforganisatie, wekken de inherente dynamische attractoren de suggestie van doelgerichtheid. Maar daar is niets teleologisch aan. De nieuwe fase in ontwikkeling wordt gekenmerkt door emergentie maar is geheel verklaarbaar uit causale factoren. Iedereen kan bij het Santa Fe Institute een gratis cursus volgen om kennis te maken met de voor evolutiebiologen onmisbare complexe adaptieve systemen.*
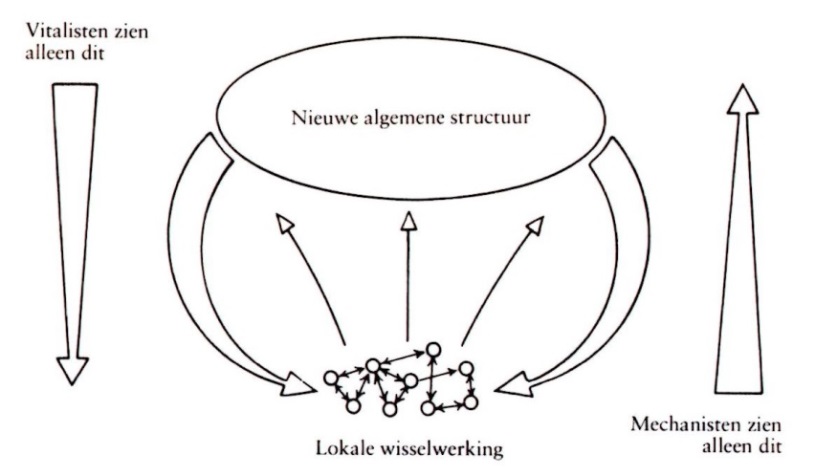
Ontstaan van nieuwe structuur (met emergente eigenschappen) in complex systeem.
uit: R. Lewin Complexiteit Contact,1993
Primordiale overgangen
De organische evolutie op aarde werd voorafgegaan door een veel langere periode van meer dan 10 miljard jaar anorganische evolutie waarin ons universum zich ontwikkelde. Astrofysische rapporten over de vroege ontwikkeling van materie en de latere oorsprong van sterrenstelsels gaan allemaal uit van een beginstadium, toen het universum volledig gevuld was met energie en materie met een oneindige dichtheid en lichte kwantumoscillaties vanwege het onzekerheidsprincipe van Heisenberg (dit laatste veroorzaakte lokale dichtheden in een verdere homogene ruimte). Vanaf het eerste moment breidde een immens heet universum zich met grote snelheid uit terwijl de aanvankelijke losse subatomaire deeltjes na verloop van tijd samenklonterden. De oorspronkelijke nucleosynthese zorgde voor de initiële synergetische stap bij de vorming van de materie. Up- en down-quarks verenigden zich om binnen een fractie van een seconde protonen en neutronen te vormen en na ongeveer driekwart van de eerste miljoen jaar waren de meeste vrije elektronen aangetrokken om atomen samen te stellen (waterstof, deuterium, helium en lithium). Vanaf dat moment werd het universum transparant.
In de lokaal dichtere gebieden van het steeds groter wordende heelal ontstonden gaswolken van lichte elementen. Binnen deze wolken werd de dichtheid zo hoog (door zwaartekracht) dat door fusie van de lichte elementen sterren en zelfs zwarte gaten ontstonden. Te grote sterren explodeerden en slingerden zware verbrandingsproducten de ruimte in. Dit proces werd geregeld herhaald, waardoor stofwolken van elementen (waaronder zwaardere) die rond een ster circuleerden, konden evolueren tot planeten, terwijl het sterk geëxpandeerde universum tot nog maar enkele graden Kelvin was gedaald.
Entropie kan worden gedefinieerd als het totaal van mogelijke arrangementen van componenten binnen een systeem, zonder het systeem als geheel te veranderen. Het is een maat voor de hoeveelheid chaos in een fysiek systeem. Volgens de tweede wet van de thermodynamica zal de entropie van een geïsoleerd systeem dat niet in evenwicht is, in de loop van de tijd toenemen en bij evenwicht een maximale waarde benaderen. Sinds het beginstadium lijkt het universum echter niet in staat te zijn om een toestand van energie-evenwicht te herwinnen omdat de expansie voortdurend versnelt. Men veronderstelt dat het universum nauwelijks zou kunnen uitbreiden als het niet geïsoleerd was van aangrenzende systemen. Toch raakt het systeem steeds verder verwijderd van een evenwichtsverdeling van energie en deeltjes naarmate de universele expansie blijft versnellen, waardoor tegelijkertijd de drang naar evenwicht wordt vergroot.
Hoe dan ook, de synergetische assemblage van elementen uit subatomaire deeltjes in de condenserende intergalactische wolken werd voortgezet zodat er steeds meer sterren werden gevormd. Volgens de accretietheorie zouden planeten zijn geëvolueerd uit het resterende circulerende stof. Ondertussen waren voldoende afgekoelde atomen in staat om moleculen te vormen. Deze formatie was een voorwaarde om verdere ontwikkeling overal in de ruimte te mogelijk te maken. Aangezien het grootste deel van het universum voor elke waarneming verborgen blijft en zelfs in ons eigen melkwegstelsel deze ontwikkeling buiten onze waarneming valt, kunnen we maar beter dicht bij huis blijven.
Op onze aarde waren de meest voorkomende elementen ijzer en stikstof. Een zwaar element als ijzer werd naar het centrum getrokken, terwijl de atmosfeer voornamelijk werd gevormd door gassen die ontsnapten uit de afkoeling van rotsachtige materialen. De aanwezigheid van zuurstof werd onmiddellijk ongedaan gemaakt door oxidatie van andere elementen zoals waterstof, koolstof en silicium, waarbij water, CO2 en kwarts ontstonden. Naast stikstof moet de vroege atmosfeer hoge concentraties kooldioxide en waterdamp (H2O) hebben bevat, evenals ammoniak, waterstofsulfide en methaan.
Niet alleen organismen zijn het resultaat van emergente evolutie, ook de niet-levende materie is er door gevormd. Dit biedt kansen voor interdisciplinaire samenhang met andere beta-vakken. De grote moleculen aan het begin van de abiogenese (RNA, eiwitten, lipiden) zijn opgebouwd uit kleinere verbindingen met waterstof (meest voorkomende stof in het heelal) en oxidatieproducten (kiezel, water, koolzuur, allerlei ‘vervuiling’). Ongeacht of de abiogenese op aarde of op een meteoriet plaatsvond, de ‘grote’ atomaire bouwstenen (C, O, N, S, F, enz.) waaruit de moleculen bestaan zijn altijd afkomstig uit interstellaire gaswolken. Door de explosieve kracht van supernova’s werden ze gevormd uit waterstof en helium, de stoffen waaraan sterren hun bestaan ontlenen (waterstoffusie).
Aanvankelijk, miljarden jaren geleden, was het heelal geheel gevuld met waterstofatomen. Door onderlinge aantrekking vormden ze clusters die door de gravitatie steeds zwaarder en heter werden (stervorming). De waterstofatomen zelf zijn in de eerste miljoen jaar na de oerknal ontstaan doordat vrije elektronen gaandeweg permanent werden ‘ingevangen’ door losse kernen (door de duurzame ontkoppeling van fotonen werd het heelal toen transparant). De atoomkernen werden enkele minuten na de oerknal geboren uit samenklittende protonen en neutronen die niet langer door energierijke deeltjes uit elkaar werden geslagen. Op hun beurt werden de materiedeeltjes (hadronen) in de eerste seconde gevormd uit samensmeltende quarks onder invloed van de sterke wisselwerking (gluonen). Eerdere ontwikkelingen in de ontzaglijke hete en dichte Oersoep is onderwerp van theoretische speculatie, het noodzakelijke energieniveau kan in geen enkele deeltjesversneller op aarde worden bereikt.
Het ontstaansproces van de materie mag dan buiten het examenprogramma vallen, als onderdeel van ‘big science’ en het emergentie paradigma mag het niet ontbreken.
Vanwege wiskundige problemen werden concepten als antideeltjes en denkbeeldige tijd geïntroduceerd. Die had men liever dan een oorzaak in de toekomst zoals sommigen voorstelden. De steeds groeiende complexiteit op microniveau leek te worden veroorzaakt door een attractor die de tweede hoofdwet van de thermodynamica tegenspreekt. Deze negentropie of syntropie wordt wel beschouwd als een analogie van antimaterie en is wiskundig afgeleid van de vierkantswortel in de energie / momentum / massa-vergelijking die zowel een positieve (convergente) als negatieve (divergente) oplossing heeft. Deze syntropie is een intrigerend concept dat complexiteit kan veroorzaken via vreemde attractoren zoals in niet-lineaire dynamische systemen. Het concept wordt echter van nature omarmd door creationisten, waardoor het gebruik ervan ongemakkelijk wordt voor wetenschappelijke toepassing. De tweede wet van de thermodynamica is alleen van toepassing op geïsoleerde systemen, dus - tenzij een steeds groter wordend universum niet als een geïsoleerd systeem wordt beschouwd - is het gebruik van syntropie beperkt tot 'open' (energie-verwerkende) systemen, zoals levende organismen.
Autokatalyse
De organische evolutie op aarde kan worden beschouwd als een voortzetting van het thermodynamische principe van energie-transductie. Vanwege het onzekerheidsprincipe van Heisenberg en het uitsluitingsprincipe van Pauli, zou een hele reeks moleculen met hun substantiële kenmerken kunnen worden geproduceerd door louter grootte-variatie van de verschillende atomen en hun onderlinge elektronenuitwisseling. Een enorme verscheidenheid aan 'vrije' chemische verbindingen (tegenwoordig zijn ze meestal verankerd in organische structuur of proces) vergroot de kans op de aanwezigheid van katalysatoren die nog meer chemische structuren mogelijk kunnen maken. In een willekeurig mengsel van reagentia wordt vroeg of laat een katalysator gesynthetiseerd. Deze katalysator zal de verdere processen binnen het mengsel in een bepaalde richting leiden door willekeurige reacties tegen te gaan. Door autokatalytische en oscillerende reacties op gang te brengen, ontstaat er een onvermijdelijk proces.
Een reconstructie van de oorsprong van het leven is hoogst hypothetisch, maar er zijn enkele onbetwistbare premissen. Om een entiteit als een levend wezen te definiëren, moet het aan ten minste twee voorwaarden voldoen. De entiteit moet i) repliceerbaar zijn om informatie over zichzelf door te geven, inclusief het vermogen om te repliceren, en de entiteit moet los van de buitenwereld ii) beschikken over een interne omgeving om energie op te wekken. Uiteindelijk kon er homeostase ontstaan - voor sommigen kan dit zelfs een voorafgaande toestand zijn - door selectieve transudatie van ionen terwijl de nodige energie werd geleverd door chemische modulaties in organisch materiaal. Dat laatste was overvloedig aanwezig door abiotische synthese in de buurt van vulkaanuitbarstingen en wellicht door elektrische ontladingen in de reducerende gasatmosfeer.
Tijdens de enorme tijdspanne dat leven kon ontstaan hoefde replicatie met waarschijnlijk een stof als het huidige RNA maar één keer te gebeuren. Door zichzelf in stand te houden zou het blijven repliceren en verdere complexiteit ontwikkelen om zo zichzelf te verbeteren. Dit zou kunnen verklaren waarom alle moderne organismen dezelfde structuren hebben om informatie over te dragen.
Na de ontdekking van katalytische RNA's (introns) bij zeer uiteenlopende organismen, is er veel gespeculeerd over hun betekenis in de vroege evolutie van het leven.
Een autokatalytisch chemisch netwerk lijkt een serieuze en plausibele kandidaat om de eerste RNA's te hebben geproduceerd. Dit kan voorafgegaan zijn door een reducerend chemisch reactie-netwerk om energie te leveren (zoals in moderne chemotrofe bacteriën). Als prebiotisch metabolisme werd een omgeving voorgesteld met autokatalytische niet-polymere moleculen, zoals moderne co-enzymen, als voorlopers van nucleotiden. Deze moleculen koloniseren het oppervlak van zelfassemblerende olie-druppeltjes, oxideren de koolwaterstoffen in de olie en veranderen langzaam maar zeker de structuur in een membraan. De verwijdering van een interne omgeving zou mogelijk ook geleverd kunnen worden door zelfgeorganiseerde lipidemembranen.
Het idee van zelforganiserende autokatalytische chemische systemen als basis voor de vorming van vroege zichzelf onderhoudende pre-metabolische moleculaire netwerken wordt serieus onderzocht en is geschikt gebleken om informatieoverdracht van de ene generatie naar de volgende mogelijk te maken.
De oeratmosfeer was sterk reducerend qua samenstelling (voornamelijk H2O, CO2 en N2) en kwam vooral voort uit ontgassing tijdens meteorietinslagen (in plaats van uit de aardmantel zelf). In tegenstelling tot wat lang werd gedacht, namelijk dat NH3 en CH4 belangrijke componenten waren van de vroege atmosfeer, zouden deze gassen alleen aanwezig zijn geweest in de buurt van vulkaanuitbarstingen en opgelost in de oceanen. De samenstelling van de atmosfeer was bepalend voor het ontstaan van leven wat betreft de aanwezige componenten in potentiële autokatalytische netwerken en met het oog op de mogelijke fotochemische reacties en het soort doorgelaten straling vanuit de ruimte. De afwezigheid van ammoniak en methaan in de atmosfeer gaf aanleiding tot de hydrothermische hypothese die een onderzeese oorsprong van leven suggereert. En er is de theorie van panspermie die veronderstelt dat biochemische verbindingen uit de ruimte afkomstig zijn. Hoewel een exogene bron van organische moleculen als oorsprong van het leven in tegenspraak is met zowel de filosofische afwijzing door Ockham's scheermes als met de meer pragmatische overweging dat slechts een kleine fractie een inslag op aarde zou overleven, wint het aan waarschijnlijkheid in een atmosfeer die wordt gegenereerd door vrijkomende gassen bij meteorietinslagen. Volgens de opvatting dat evolutionaire veranderingen met hele kleine stapjes heeft plaats gevonden, zou het leven lang voordat de aarde werd gevormd zijn begonnen en daarom op een andere planeet moeten zijn ontstaan. De berekeningen kunnen echter op verkeerde aannames gebaseerd zijn en onvoldoende rekening houden met de attractoren in de dynamische subsystemen, wat de accumulatieperiode aanzienlijk zou kunnen verkorten.
Van de talrijke microsferen moeten sommige een replicerende structuur hebben omsloten. De kans op een goede combinatie was misschien miniem, het aantal moleculen en de beschikbare tijd waren daarentegen immens. Punt is dat de eerste complexe structuren die aan de meest fundamentele eisen van levensvatbaarheid voldeden, evolueerden door fusie van de juiste ingrediënten in plaats van door selectie. De eigenschappen van de losse componenten bleven intact en hun combinatie leverde iets op dat er nooit eerder was. Perfecte synergie. Of liever, emergentie.
Vroege organische evolutie
Neem een enkele oude microbe die zich slechts één keer per jaar repliceert (als de omgeving optimaal is, kunnen moderne microben zich elke 20 minuten delen) en stel dat de diameter ervan 1μm bedraagt, dan zou na het totale wateroppervlak op aarde bedekt na 100 jaar bedekt zijn met een bacteriële deken van meer dan 3μm dik. Je kunt dit zelf berekenen voor 2100 microben met een afmeting van 1 x 1 x 1 μm over een oppervlak van 3,7 x 108 km2.
De eerste protobionten (een combinatie van een omhullend membraan, een replicatieapparaat en voldoende katalytisch vermogen om de motor te laten draaien) waren hoogstwaarschijnlijk chemotrofen met een omgekeerde Krebs-cyclus die al het beschikbare (an)organische materiaal 'consumeerden'. De energie die door deze autotrofe replicatoren werd gegenereerd, zou afval produceren dat vroeg of laat op een vergelijkbare manier door andere protobionten kon worden gebruikt. En hun afval weer door anderen, enz. Het is het begin van een organische co-evolutie (onderling afhankelijke relatie) en tegelijkertijd zou natuurlijke selectie (competitie) zijn biologische debuut maken. Autokatalytische replicatie van losse bouwstenen werd gereguleerd (we weten nog niet hoe) en omgezet in reproductie.
De oersoep van protobionten zag er misschien wat chaotisch uit met veel interacties en in- en uitsluitingen van grotere en minuscule bio-druppeltjes. Het minimaliseren van het aantal membraanverbindingen resulteerde in een bolvorm vanwege de fysische wetmatigheden. Chemische eigenschappen van de verbindingen konden leiden tot plooien en uitrekking als de omgevingsdruk hoger was dan de morfogenetische kosten, zo die er al was.
De vroegste prokaryote fossielen - tegenwoordig wordt de term prokaryoot vervangen door echte bacteriële - dateren van ongeveer 3,8 miljard jaar geleden (nabij Quebec). Aangezien de aarde zelf hooguit 4,5 miljard jaar oud is, moeten deze prokaryoten gedurende de eerste paar honderd miljoen jaar zijn geëvolueerd - de suggestie dat levende organismen de aarde vanuit de ruimte hebben gekoloniseerd leidt alleen maar af en maakt de zaak onnodig ingewikkeld. Het lijkt aannemelijk dat veel van de oorspronkelijke levensvormen zijn verdwenen nadat ze werden opgeslokt, opgenomen of ingekapseld door meer succesvolle vormen die in staat waren om specifiek organisch materiaal op een meer efficiënte wijze te verwerken. Dit zou natuurlijke selectie kunnen worden genoemd, maar niet zoals Darwin het definieerde. Eén oude groep bacteriën, de Archeae, bestaat echter nog steeds en ze onderscheiden zich van alle andere moderne levensvormen door een specifieke samenstelling van hun membranen. Tot de Archeae behoren de enige moderne levensvormen die extreme temperaturen en zoutconcentraties (extremofielen) kunnen overleven.
Moderne bacteriën zijn enorm talrijk en kunnen op verschillende manieren worden geclassificeerd. Het meest algemeen zijn de fotolithotrofen die licht en een anorganische elektronendonor (H2O, H2S of S) gebruiken en de chemoorganotrofen die redoxreacties en een organische elektronendonor gebruiken. Om aan koolstof te komen nemen de autotrofen zoals cyanobacteriën CO2 op of breken ze organisch materiaal af (heterotrofen zoals parasieten en reducenten). Aërobe hetrotrofen gebruiken zuurstof als elektronen-acceptor, anaërobe geven hun elektronen af aan een verscheidenheid van (an)organische chemicaliën. Sommige van de aanwezige bacteriën zijn aëroob maar niet autotroof (nitrificatiebacteriën), andere gebruiken licht maar geen CO2 (paarse bacteriën). De immense diversiteit van tegenwoordig moet destijds beslist onbeduidend zijn geweest aangezien moderne bacteriën in hoge mate afhankelijk zijn van andere levensvormen (en vice versa). Toch vormden ze een onvermijdelijk matrix voor verdere organische evolutie.
Reproductie van bacteriën wordt voornamelijk gerealiseerd door binaire splitsing, terwijl verschillende manieren van recombinatie voorhanden zijn. De onverwachte, vaak voorkomende laterale overdracht van genetisch materiaal tussen verschillende bacterie-stammen wil niet zeggen dat deze ook drie miljard jaar geleden voorkwam, maar ondersteunt nog wel steeds het idee van emergente evolutie.
Ervan uitgaande dat er voldoende organisch materiaal beschikbaar was voor een wereldwijde groei, lijdt het geen twijfel dat, naast selectie door genetische mutaties, de ophoping van microben gemakkelijk allerlei soorten fusies tot gevolg had. Een direct bewijs voor emergente evolutie tijdens de eerste miljard jaar van organische evolutie is niet voorhanden. Het is wel aannemelijk.
Endosymbiose
De echte bacteriën hebben celmembranen die vergelijkbaar zijn met die in alle andere organismen (behalve de Archeae). Dit feit, evenals hun eenvoudige structuur (geen organellen; vrij DNA), maakt het aannemelijk dat alle eukaryoten zijn geëvolueerd uit de echte bacteriën. Onder de vele complexe moleculen waren er enkele straling-absorberende eiwitten, via welke elektronen op andere moleculen konden worden overgebracht om daarmee wat energie op te wekken. Een primitieve fotosynthese verving bij sommige bacteriën de chemische omzettingen die nodig waren om energie op te wekken. Het bijproduct zuurstof kan voor veel van de organische structuren behoorlijk schadelijk zijn geweest.
De opmerkelijke overeenkomsten tussen vrijlevende cyanobacteriën en de chloroplasten in moderne plantencellen waren een eerste aanleiding tot de ontwikkeling van de seriële endosymbiotische theorie. Hoewel niet unaniem gesteund wordt nu algemeen aangenomen dat zowel chloroplasten als mitochondriën zijn geëvolueerd uit bacteriën die ingekapseld werden door een grotere entiteit. De combinatie van fotosynthese en zuurstofrecycling was een volgende stap in de emergente evolutie. De basissubstantie van de eukaryote cel, het nucleocytoplasma, is mogelijk ontstaan uit een associatie tussen archeae en anaërobe heterotrofe bacteriën, terwijl het nucleaire omhulsel en het endoplasmatisch reticulum tot stand kwamen door zelfassemblage. Deze zogenaamde proto-eukaryoten zouden dan cyanobacteriën en paarse niet-zwavelbacteriën hebben opgenomen die we nu kennen als resp. chloroplasten en mitochondriën. Deze hypothese over een ontwikkeling van enkele miljarden jaren geleden verdient enig voorbehoud en moet misschien op details worden herzien. Er bestaan tegenstrijdige theorieën over de oorsprong van trilharen en andere bewegingsstructuren in eukaryoten. Voorstanders van de seriële endosymbiotische theorie beweren dat voorouders van moderne spirocheten zijn doorgedrongen in de proto-eukaryoten om microtubuli en undulipodia te vormen. Endogene modellen gaan ervan uit dat de flagella zijn ontstaan uit componenten van het eukaryote cytoskelet.
Ook over de oorsprong van de eukaryote kern zijn de meningen verdeeld. Het endogene model is gebaseerd op de aanname dat de kern selectief is voortgekomen uit een gemodificeerd reeds bestaand endomembraansysteem. Volgens het syntrofische model is de kern ontstaan uit een fusie tussen archaea en bacteriën of tenminste uit de endosymbiotische opname van archeaeïsche eigenschappen in een gastcel. Er is ook toenemende belangstelling voor de mogelijke rol van virussen bij het ontstaan van de eukaryoten.
Van eencellig naar meercellig
Gedeeltelijke mitose, aan elkaar blijven plakken na deling, kolonievorming, er zijn diverse mogelijkheden hoe meercellige organismen uit enkele cellen zijn voortgekomen. Wat ze gemeen hebben, is de noodzakelijke samenwerking tussen de cellen. Sommigen denken dat het ontstaan van meercellige organismen een natuurlijk gevolg was van een kolonie cellen die continu kon groeien. Alle water-organismen werden meercellig door gedeeltelijke mitose terwijl bij de meeste terrestrische micro-organismen sprake was van een soort beweeglijke aggregatie van cellen of kernen in een meerkernig syncytium. Of de meercellige organismen voortkwamen uit aggregaties van gelijkaardige eencellige organismen zoals in de huidige slijmzwammen of dat multicellulariteit voortkwam uit verschillende soorten eencellige organismen (symbiose-theorie), in beide gevallen zou men kunnen spreken van emergente evolutie. In nauw verwante cellen kan hun samenwerking worden begrepen uit het oogpunt van 'egoïstische genen' terwijl de vorming van kolonies van minder verwante individuele cellen verklaard zou kunnen worden met het 'prisoners dilemma'. Aangezien multicellulariteit waarschijnlijk vele malen onafhankelijk is geëvolueerd, zijn mogelijk alle scenario's gerealiseerd.
Onlosmakelijk verbonden met multicellulariteit zijn celdifferentiatie en apoptose (celdood) die mogelijk hun oorsprong hebben in hun eigen geschiedenis, dat wil zeggen, de eencellige evolutie. De toegenomen genoomcomplexiteit die samenvalt met multicellulariteit houdt hoogstwaarschijnlijk verband met intercellulaire communicatie en eerder genoemde processen.
Samenwerkende eenheden van cellen met verschillende taken zullen op het terrein van vruchtbaarheid en levensvatbaarheid een enorm voordeel hebben opgeleverd ten opzichte van eencellige organismen. Bij eencellige individuen moet dezelfde cel voor beide fitnesscomponenten zorgen, terwijl bij meercellige organismen afzonderlijke cellen zich kunnen specialiseren. Selectieve druk is voordelig bij grote verscheidenheid en kan eencellige organismen aanzetten tot kolonievorming om vervolgens te evolueren tot meercellige individuen. Grootte kan gunstig zijn voor de levensvatbaarheid (zoals predatie vermijden, grotere prooien vangen, buffervorming) en voor de vruchtbaarheid. Naarmate de grootte verder toeneemt, kunnen de voordelen voor levensvatbaarheid en vruchtbaarheid nog beter worden door kiemvorming en de volledige scheiding tussen kiem en soma. Met het resultaat wordt een verhoogde complexiteit bereikt.
Toen meercellige organismen ruim een half miljard jaar geleden ontstonden, waren alle fundamenteel verschillende bouwplannen van de moderne phyla al vertegenwoordigd. De variatie in bouwplan bleef beperkt door intrinsieke wetmatigheden van de sub-systemen tijdens de ontogenie. Ongeveer 50 miljoen jaar later verdween 85% van de meercellige organismen en verdwenen de meeste phyla voor altijd. Deze eerste bekende massa-extinctie viel samen met de vorming van Gondwanaland. Blijkbaar werkten de externe omstandigheden (verkorte kustlijn en verlaagde gemiddelde temperatuur) als een selectief mechanisme ten gunste van slechts enkele bouwplannen.
Volgens het fossielenbestand kwamen na de multicellulariteit de terrestrische planten en dieren relatief snel tevoorschijn. Aangezien de oudste overblijfselen van organisch aards leven meer dan 2 miljard jaar oud zijn, is het aannemelijk dat het meeste ervan destijds beschikbaar was als voeding voor de eukaryote nieuwkomers. Algemeen wordt aangenomen dat de eerste terrestrische eukaryoten autotroof waren, zodat ze bijdroegen aan een verdere toename van het zuurstofgehalte in de atmosfeer. Omdat de ozonlaag nog niet volledig was ontwikkeld was het stralingsniveau bedreigend hoog. Daardoor was de aarde voor de nieuwe levensvormen een levensgevaarlijke omgeving. Het land was voedselarm, fysiek onstabiel en onderhevig aan sterke temperatuurschommelingen. Alleen alluviale of lacustriene laaglandgebieden met fijnkorrelige bodems die water en voedingsstoffen konden vasthouden, waren mogelijk een geschikte habitat voor de eerste landplanten.
Men vermoedt dat de kolonisatie van het land door eukaryoten waarschijnlijk werd vergemakkelijkt door symbiose tussen een fototroof en heterotroof organisme. Het fossielenbestand in dit opzicht is ontoereikend. De symbiotische samenwerking tussen schimmels en algen of cyanobacteriën bestond echter al lang vóór de evolutie van vaatplanten en hun synergetisch aandeel in de terrestrische invasie was onvermijdelijk. Met hun hyfen konden de schimmels water en opgeloste anorganische ingrediënten opnemen, terwijl de ingesloten fototrofen de benodigde suikers leverden. Fotosynthetiserende algen waren veel te kwetsbaar om alléén te overleven en werden beschermd tegen te grote hoeveelheden UV-straling en uitdroging door de cuticula van de schimmel.
De meeste moderne embryofyten (landplanten) onderhouden nog steeds een symbiotische relatie met schimmelhyfen in de vorm van mycorrhiza, om het contactoppervlak van het wortelsysteem van de plant te vergroten. Bovendien kan de mycorrhiza voedingsstoffen leveren die zonder deze samenwerking voor de plant niet beschikbaar zouden zijn.
Moderne korstmossen bestaan uit een symbiotische associatie van een schimmel en een groene alg. Ze kunnen extreme omstandigheden overleven, hoewel sommige soorten behoorlijk gevoelig zijn voor luchtverontreiniging. Uit onderzoek blijkt dat daarnaast ook bacteriën verschillende functionele rollen kunnen vervullen binnen de symbiose van korstmossen.
Verschillende moderne groepen kolonievormende algen zijn tussenproducten tussen eencellige en meercellige planten. Het meest bekend onder de groene algen zijn de bolvormige kolonies van Volvocales die zijn samengesteld uit talrijke flagellaten met af en toe cytoplasmatische verbindingen en een duidelijke voor- en achterzijde. Groene algen van het geslacht Scenedesmus scheiden prikkels af om kleine niet-beweeglijke kolonies te vormen, waarschijnlijk om predatie te voorkomen (de kolonie is te groot om door zoöplankton te worden geconsumeerd). Veel goud- en geelbruine algen (chrysophyta) en diatomeeën (bacillarioophyceae) kleven aaneen tot kolonies, de laatste met een rudimentaire celdifferentiatie.
Moderne slijmzwammen beginnen als een eenkernige amoebe. Tijdens de groei delen alleen de kernen zich en ontwikkelt het organisme zich tot een groot meerkernig plasmodium. Als de omstandigheden gunstig zijn voor vruchtvorming, valt het plasmodium uiteen om een vruchtlichaam te vormen dat haploïde sporen draagt. Daaruit ontwikkelen zich de gameten van de volgende generatie.
De moderne porifera (sponzen) nemen als meercellige organismen een positie in tussen de eencellige protozoa en de metazoa met een zenuwstelsel. Van alle moderne dieren zijn ze het nauwst verwant met de voorouderlijke oermetazoa. Ze zijn samengesteld uit een beperkt aantal gedifferentieerde cellen die, na scheiding, weer samenklitten tot het oorspronkelijke dier. Het bestaan van cel-adhesiemoleculen maakte de ontwikkeling van een koloniaal organisme mogelijk. Sponzen hebben geen zenuw- en spierweefsel. Cellulaire communicatie en coördinatie worden chemisch gegenereerd door een complexe reeks moleculaire componenten in plaats van door elektrische geleiding.
UITSTERVEN
Gedurende vier miljard jaar aan levensgeschiedenis op aarde zijn er regelmatig massale uitstervingen geweest waarbij de dominante soorten het veld moesten ruimen en plaats maakten voor nieuwe. Waardoor de overheersers verdwenen is vaak ongewis en naarmate de gebeurtenissen zich recenter afspeelden is zijn er meer details bekend. Zoals het massale sterven van veel soorten in het Meghalayaan (de afgelopen 4500 jaar) a.g.v. ziekteverwekkers, vervuiling en verdringing door Homo sapiens. Maar de alomvattende overheersing van de mens heeft tijdens het anthropoceen nog niet tot massaal uitsterven van andere soorten geleid zoals in het verleden.
Uitstervingsgolven zijn essentieel voor de evolutie, ze bieden ruimte voor de ontwikkeling van nieuwe soorten. Er verdwijnt in een relatief korte tijd minstens 75% van de voorkomende soorten.
Sinds het bestaan van meercellige organismen (planten, dieren en schimmels) hebben er zes grote catastrofen plaatsgevonden. De eerste was in het Ordovicium, ca. 450 miljoen jaar geleden (450 Ma) en reduceerde het aantal soorten met 85%. Een sterke daling van het CO2 gehalte in de atmosfeer, mogelijk door een ongebreidelde toename van landplanten, leidde tot een ijstijd. Verandering temperatuur en zeeniveau waren de boosdoeners maar de oorzaak is onbekend. De tweede uitstervingsgolf vond plaats in het Devoon, ca. 400 Ma en werd mogelijk veroorzaakt door meteorietinslagen. Ook deze periode wordt gekenmerkt door een sterke fluctuatie van het CO2 gehalte. De derde en vierde golf vonden betrekkelijk kort na elkaar plaats, resp. 260 en 250 Ma. Daarbij verdwenen bijna alle soorten van de aardbodem. De oorzaak wordt gezocht in wijdverbreid vulkanisme (vloed-basalten) waardoor het CO2 gehalte steeg, met sterke temperatuurstijging en verzuring van de oceanen tot gevolg. De volgende massa-extinctie, ca. 200 Ma, treft vooral het landleven waar ca. 80% van de soorten verdwijnen. De oorzaak is onbekend maar ook hier vermoedt men vulkanisme en klimaatverandering. De laatste uitstervingsgolf vond ca. 65 Ma plaats door de inslag van een komeet in Yucatan, waarbij 50% van het zeeleven en alle grote landdieren het loodje legden.
De oudste bekende massa-extinctie was de grote oxygenatie, ca. 2500 Ma, toen cyanobacteriën met fotosynthese grote hoeveelheden zuurstof produceerden. Oxidatie bedreigde alle anaerobe levensvormen en tastte de broeikasgassen aan, met een wereldwijde afkoeling tot gevolg, die bijna fataal was voor de cyano-bacteriën zelf. De eerste meercellige organismen waren sessiel en verdwenen ca. 540 Ma massaal, kort voor de Cambrische explosie. Mogelijk werden ze slachtoffer van predatie door organismen die zich wel konden voortbewegen.
Heel recent, ca. 2 Ma, zou een reeks supernova’s de ozonlaag hebben aangetast waardoor een groot aantal kleine organismen door kosmische straling werd gedood. Hierdoor werd het ecosysteem van de oceanen verstoord. In hoeverre dat van invloed is geweest op de concentratie broeikasgassen en de afwisselende ijstijden en interglacialen is onbekend.
Massa-extincties zijn van invloed op het verloop van de evolutie. De samenhang met emergentie is discutabel.*
Op de geologische tijdschaal bestaan wij nog maar betrekkelijk kort, hooguit 5 miljoen jaar. Toen liepen er primaten rond met woekerende zenuwcellen in hun hersenpan en een heterochronische verandering in hun ontwikkeling. Deze afwijking veroorzaakte neotenie (vroeggeboorte) wat goed van pas kwam omdat de uitdijende schedel nog maar nauwelijks het geboortekanaal kon passeren (een te grote schedel heeft wellicht bijgedragen aan het uitsterven van sommige Hominidae). De vertraagde ontwikkeling door heterochronie leidde ook tot uitstel van de secundaire geslachtskenmerken en tot een toegenomen afhankelijkheid van soortgenoten. De langdurige zorg die de jongen vereisten stimuleerde het sociale gedrag van deze groepsdieren.
Van de Hominidae bestaan er nog 8 soorten, de grote mensapen en wijzelf. De rest, een kleine 50 soorten, is uitgestorven. Dat Homo sapiens de vroege strijd om het bestaan heeft kunnen overleven wordt voornamelijk toegeschreven aan zijn cognitieve en coöperatieve vermogens. Een sterke neiging tot zelfopoffering, vooral ten aanzien van de nakomelingen, moet niet worden uitgesloten. Onderlinge afhankelijkheid door individuele kwetsbaarheid heeft wellicht vroege populaties helpen overleven. Maar het blijft verrassend.*
Sociaal gedrag en symbiose
Het optreden van symbiose, als een onderling afhankelijke samenwerking tussen verschillende soorten, is hierboven al genoemd als aanwijzing voor emergente evolutie. Symbiose is een bron van evolutionaire nieuwigheid en speelt een cruciale rol bij soortvorming en ecologische diversificatie. Symbiotische relaties tussen meercelligen en microben kunnen ervoor zorgen dat de meer complexe levensvormen kunnen overleven, zelfs onder extreme omstandigheden. De symbionten kunnen zodanig versmolten zijn dat ze niet zonder elkaar kunnen leven. Dit hoeft niet altijd het geval te zijn bij de wederzijds voordelige interacties tussen planten en dieren. Toch speelt het symbiotische netwerk van hun interacties een prominente rol in het functioneren van een ecologisch systeem en zorgt het zelf weer voor een hoger niveau van complexiteit. Wederom emergentie. Er zit misschien geen aanwijsbare kracht achter maar de stijgende trap van de emergente evolutie is onmiskenbaar.
Bij eukaryoten wordt het vertonen van gedrag gewoonlijk alleen aan dieren toegekend. Als de interactie tussen individuen van een soort een bepaalde mate van organisatie overstijgt kan er sprake zijn van sociaal gedrag. De definitie is vaag aangezien er veel gradaties van socialiteit zijn en vooral de hoogste niveaus (eusocialiteit) zijn het vermelden waard in het kader van de emergente evolutie.
Het ontstaan van geïndividualiseerde kolonies heeft meerdere keren in verschillende dierlijke phyla plaats gevonden. Het bekendst zijn de sociale insecten, maar eusocialiteit komt ook voor bij garnalen en molratten. Evolutionisten hebben zich lange tijd afgevraagd hoe sociaal gedrag (en altruïsme) kon evolueren. Daarbij gingen ze uit van en genetisch selectieve in plaats van een synergetische benadering. Dat leverde verwantschapsselectie en de egoïstische gentheorie. Eusocialiteit is echter het gevolg van groepsselectie in plaats van individuele selectie en kolonies zijn consequent superieur aan hun solitaire concurrenten (waaruit ze voortkomen). In plaats van moleculaire oorzaken (zoals een gen voor eusocialiteit) lijken het juist ecologische beperkingen te zijn die eusocialiteit bevorderen. De genetische verwantschap is juist een gevolg zijn van vijandelijke druk die individuen dwingt tot samenwerking bij het bouwen van hun complexe nesten en het verdedigen van hun waardevolle voedselhabitat. Hoe groter de kolonie hoe beter. Eusocialiteit lijkt vooral het resultaat van een onvoorspelbare combinatie van omstandigheden. Waardevolle voedselbronnen bevorderen de samenwerking tussen nauwe verwanten. Ze kunnen gemakkelijk door concurrenten en/of roofdieren worden veroverd wat het de moeite waard maakt om ze te verdedigen. Vijandelijke aanvallen op zulke familiecoöperaties kan bij het verdedigen van de kolonie de evolutie van de beveiligingsspecialisten bevorderen en de kans op zowel hun eigen voortbestaan als op dat van de voortplantingsspecialisten verhogen.
Bijenteelt bestond al in het oude Egypte maar pas in de jaren '40 kreeg Von Frisch inzicht in hun communicatieve kwaliteiten. Wellicht worden er in de toekomst meer verbazingwekkende manieren van communicatie bij sociale insecten ontdekt. Hoewel analogieën vaak bedrieglijk zijn is het onweerstaanbaar om communicerende mieren te vergelijken met zenuwcellen en de mieren-kolonie, als een soort superorganisme, met het zenuwstelsel.
Symbiose en eusocialiteit komen samen wanneer sociale dieren andere organismen uitbuiten. Bekend is het kweken van schimmels en bladluizen door sommige mieren. Sommige soorten gebruiken andere mierensoorten als slaven en er zijn symbiotische relaties met planten, bv. acaciabomen. Minder bekend is de relatie tussen de eusociale garnalen en koraalrifsponzen. Interessant is de symbiotische relatie tussen kakkerlakken en interne microben waarvan wordt beweerd dat ze eusocialiteit hebben veroorzaakt bij termieten. De wederzijdse afhankelijkheid tussen totaal verschillende organismen, waarvan er zelfs één met zijn eigen complexe eusocialiteit, geeft inzage in een aards ecosysteem, wellicht zelfs inzage in de gehele biosfeer, die doet denken aan de microbenmatten die ooit de aarde bedekten, maar dan zo veel complexer. Het is niet verrassend dat deze context van op elkaar inwerkende onderling afhankelijke levende wezens ooit werd opgevat als een autopoietisch systeem en anderen inspireerde tot het formuleren van de Gaia-hypothese.
Sommige dieren leven onder moeilijke omstandigheden in symbiose met soorten uit andere hoofdgroepen. Chemoautotrofe bacteriën hebben een symbiotische relatie met de gigantische buisworm Riftia pachyptila . Door oxydatie van H2S van de 'zwarte rokers' op de oceaanbodem langs de Galapagos-kloof, voorzien deze bacteriën de wormen van de nodige energie om organische verbindingen te synthetiseren. De details van het symbiotisch proces zijn nog onduidelijk. Andere organismen, zoals mosselen, gebruiken vergelijkbare symbiotische mechanismen om aan hun voeding te komen. Andere dieren onderhouden een symbiotische relatie met algen die zuurstof leveren door fotosynthese in ruil voor afbraakproducten (stikstof). De zeeslak Elysia chlorotica heeft is zelf in staat tot fotosynthese door chloroplasten van algen op te nemen in de epitheelcellen van zijn spijsverteringskanaal die bovendien essentiële plastide-eiwitten leveren door horizontale genoverdracht.
De complete verovering van het luchtruim zou op termijn een andere evolutionaire uitdaging kunnen zijn. Ongetwijfeld zit de lucht al vol met kleine organismen, voornamelijk bacteriën. Alleen onder de insecten en vogels zijn hele goede vliegers te vinden. Insecten kunnen pas vliegen als de laatste fase van de metamorfose is aangebroken, dat wil zeggen, als ze volwassen zijn. Het geeft ze meer gelegenheid een partner te vinden en zich voort te planten. Eten doen ze tijdens de eerdere stadia van hun groei. Deze dieren blijven afhankelijkheid van op de grond aanwezige voedingsstoffen, zoals bijvoorbeeld bladeren, bloed of nectar. Veel vogels hebben dat probleem niet omdat ze zich voeden met vliegende insecten. Thans zijn er verschillende soorten vogels, zoals de snelle gierzwaluw, die maanden in de lucht kan blijven en zelfs slaapt tijdens het vliegen. Na tig eeuwen zijn deze vogels wellicht volledig onafhankelijk van het aardoppervlak, in die zin dat ze tijdens het vliegen hun eieren en jongen met zich meedragen. De kans van slagen zit hem vooral in de bereidheid tot samenwerking, in dit geval met soortgenoten.

Wereldwijde netwerken
Ondanks ons relatief korte bestaan op aarde hebben we het gevoel dat die onvoorstelbaar lange evolutie er niet zo veel toe doet. Het wil niet echt tot ons doordringen dat we zonder archaea en bacteriële microben niet zouden bestaan. Ongeveer zoals een blaadje in een boomkruin zonder die boom er ook nooit zou zijn geweest. Wij bestaan door alles wat ooit eerder is geweest.
Onze hersenen zijn niets meer dan een proliferatie van zenuwweefsel. We ervaren ons brein echter als de zetel van onze cognitie en (zelf)bewustzijn, van onze ziel zelfs (wat dat ook mag wezen; misschien is het niets anders dan een soort spinoff). Het grootste deel van onze hersenen is terug te vinden in andere gewervelde dieren, om waar te nemen en te reflecteren op gevoelens. Bovendien worden onze grote frontale kwabben voornamelijk gebruikt bij het verwerken van gegevens, een eigenschap die zich alleen zo goed heeft kunnen ontwikkelen omdat we extreem sociaal wezens zijn. Die enorme capaciteit om data te verwerken heeft het mogelijk gemaakt om een belangrijke innovatie te ontwikkelen: de ongeëvenaarde manipulatie van de minerale wereld die een onvoorziene stijging op de ladder van de emergente evolutie opleverde, namelijk de ontwikkeling van ruimtevaart en van cyberspace. Critici van de Gaya-hypothese, waarin de biosfeer wordt voorgesteld als een soort superorganisme, wezen op het onvermogen van de aarde om zich voort te planten. Dit werd al in 1973 tegengesproken door Sagan die wees op onze potentie om de ruimte te verkennen en andere planeten te koloniseren.
Het wereldwijde informatienetwerk dat onze soort heeft aangelegd, zorgt voor een volgende stap in emergente evolutie. Tegenwoordig kan praktisch elk mens met iedereen op deze planeet communiceren. Binnen enkele seconden kan informatie over onderhoud of verbetering van kleinere of grotere delen van de samenleving, naar alle uithoeken van de aarde worden verspreid. Veel van die informatie is natuurlijk irrelevant en zelfs onjuist, net zoals dit het geval is in onze hersenen en ons DNA. Dat verandert echter niets aan de synergetische impact van cyberspace op het dagelijks leven.
Niet alleen informatie is belangrijk. Er is ook een geraffineerde stroom van economische afhankelijkheid die over de planeet slingert, om de nodige energie te leveren voor het behoud van sociale gemeenschappen (vaak ten koste van anderen). De belangrijkste kracht die dit proces in stand houdt, is de universele drang van kleinere eenheden, zelfs van individuen, om de eigen invloed te vergroten.
En last but not least, is er, mogelijk als essentieel onderdeel van eusocialiteit, een fundamentele menselijke behoefte aan veiligheid die alles aanstuurt, ook als we het niet met elkaar eens zijn.
Tegenwoordig is er ook veel zorg voor ons milieu. Veel mensen zijn bang dat allerlei soorten organismen uitsterven door onze groeiende dominantie. Vele van de bestaande organismen zijn als voedselbron noodzakelijk voor ons voortbestaan en andere zijn, door de gesaldeerde relaties zoals hierboven beschreven, indirect nodig voor onze geestelijke en lichamelijke gezondheid (al kunnen we sommige missen als de pest). We zijn het resultaat van een evolutionaire ontwikkeling maar dat geldt ook voor alle andere levende wezens. En veel van hen kun je eigenlijk als doodlopende wegen opvatten. De meeste mensen hebben geen probleem met het uitroeien van onkruid en ongedierte maar anderzijds worden bouwprojecten opgeschort ten behoeve van een onschadelijke maar zeldzame soort. Het lijdt geen twijfel, ons bestaan maakt louter deel uit van de wereld als geheel.
We vinden het vanzelfsprekend dat alles is opgebouwd uit atomen en moleculen en we gaan ervan uit dat deze kleine bouwstenen ook als vrije deeltjes voorkomen. Maar wij vinden het minder vanzelfsprekend dat de 1030 bacteriën op onze planeet de helft van alle biomassa op aarde vertegenwoordigen. En zonder hen zou de eukaryote cel nooit zijn geëvolueerd. Er bestaan nog steeds minstens een kwart miljoen eencellige eukaryote soorten, ondanks of wellicht dankzij het ontstaan van planten, dieren en schimmels. Niettegenstaande het grote aantal parasieten waren meercellige organismen waarschijnlijk nooit geëvolueerd zonder de reeds aanwezige bacteriën en eencellige eukaryoten. Eusocialiteit en menselijk sociaal gedrag werden eerder versterkt door de aanwezigheid van andere levensvormen (in landbouw, veeteelt en sanitaire voorzieningen). De overlevingskans van elk organisme hangt in de eerste plaats af van consumeren en vermijden zelf geconsumeerd te worden. In het kader van de grote transities lijken mensen op protobionten: allebei maken ze gebruik van levenloze materie om zich te verspreiden (door replicatie, reproductie, communicatie en buitenaardse verkenning). Het wereldwijde netwerk is niet beperkt tot een op silicium gebaseerd informatienetwerk, het is ook het totaal van gelaagde organische interacties (de bovenliggende worden gewoonlijk ecosystemen genoemd) en het is ook onderworpen aan de wetten die gelden binnen ons zonnestelsel, ons melkwegstelsel, het niet-lokale universum waarvan ze deel uitmaakt.
De belangrijkste bijdrage die het emergentie-paradigma het onderwijs biedt is de kans om op grond van het curriculum antwoord te geven op zinsvragen zonder ons in geloofskwesties te mengen (voor zover dat door evolutiedenken niet al gebeurd is). Emergente Evolutie is al omarmd door de christelijke filosofie (Klapwijk) door er Genesis met de haren bij te slepen (alsof er alleen een Bijbels scheppingsverhaal bestaat). Maar emergente evolutie heeft helemaal geen christelijke signatuur nodig om de zin van het bestaan te duiden. Om uit te leggen waarvoor wij leven, wat het doel is van ons bestaan op aarde. Het is een kwestie van extrapolatie.
De achtereenvolgende faseovergangen en organismale transities vertonen een wetmatigheid waar we niet omheen kunnen. Uit lokale wisselwerking ontstaat door zelforganisatie een nieuwe, emergente structuur. Het lijkt telkens of er een nieuw tijdperk is aangebroken, een letterlijk ongekende nieuwe wereld. De laatste fase die genoemd werd is die van de menselijke samenleving maar dat is nog niet de laatste fase van de emergente evolutie. De door de mens geïnitieerde technologische ontwikkeling is inmiddels in het stadium van de kunstmatige neuronale netwerken en evolutionaire algoritmen en schrijdt voort op het gebied van cybernetica en biomedische technologie (alweer zo’n heerlijk handvat voor disciplinaire samenhang). Het zal nog wel even duren maar een onvermijdelijk transhumanistisch tijdperk (waarover wellicht meer in een volgend artikel) ligt in het verschiet. Welke emergenties dat met zich meebrengt is per definitie ongekend.
De zin van ons bestaan is duidelijk. Wij zijn op aarde om de volgende fase mogelijk te maken. En dan heb ik het niet over het hiernamaals of het Koninkrijk Gods, al mag iedereen daar best in blijven geloven, dat doet aan de werkelijke toekomst niets af.
De toekomst
Algemeen wordt aangenomen dat evolutionaire processen min of meer willekeurig zijn. Als het proces opnieuw zou worden gestart met alle vergelijkbare omstandigheden, zou het resultaat in detail heel anders zijn. Ik ben het eens met de mening dat de oorsprong van soorten in hoge mate afhangt van toevallige omstandigheden, een natuurlijke ontwikkeling die geen herhaling toelaat. Wat betreft de primaire overgangen, ben ik echter overtuigd van een fundamentele wet waardoor overal in het tijd-ruimte continuüm, mits voldoende tijd, elegante systemen samensmelten om een nieuw niveau van complexiteit te betreden. Door deze filosofie kan ik niet alleen achteruit kijken, maar ook voorspellingen doen over de toekomst. Op aarde nam tijdens de overgang van moleculen naar het ontstaan van de mens de grootte stapsgewijs toe met een factor 109 (van 10-10 tot 10-1) over een periode van ongeveer 4,5 miljard jaar. Als we kijken naar het huidige wereldwijde netwerk is er sinds het ontstaan van de mensachtigen 5 miljoen jaar geleden een groottetoename met een factor 108 (van 10-1 tot 107). Dienovereenkomstig valt te verwachten dat aan het einde van het huidige millennium ons zonnestelsel (1012) gekoloniseerd is.
Een optimistisch beeld van de menselijke vooruitgang voorspelt een exponentiële groei van duurzame technologie en cybernetische ontwikkelingen waarmee uiteindelijk alle ecologische en sociaal-culturele problemen worden opgelost (waarschijnlijk ten koste van de massa). Het wereldwijde informatienetwerk zal met toekomstige technologische innovaties een nieuwe transitie opleveren. Homo sapiens zal de eerste impuls geven aan een nieuw soort evolutie, namelijk de ontwikkeling van zelfassemblerende robots die de melkweg gaan koloniseren.
Pessimisten, aan de andere kant, denken niet dat de menselijke hebzucht het hoofd zal kunnen bieden aan de wraak van Gaia. Zij verwachten dat de menselijke beschaving niet is opgewassen tegen de kleinschalige natuur en ze vrezen een definitief uitsterven van Homo sapiens. Als de menselijke soort inderdaad een doodlopende weg is, zal er wellicht een ander evolutionair pad worden gevolgd. Een aantrekkelijk transitie-scenario combineert gedragskenmerken (dieren) en fotosynthese (planten) om eusociale dieren te ontwikkelen die voor hun overleven niet langer afhankelijk zijn van organische verbindingen. Dit is natuurlijk fictie en het zou waarschijnlijk honderden miljoenen jaren duren voordat dergelijke organismen tot ontwikkeling zijn gekomen. Als we de zin van ons bestaan willen zien als een voorbereiding op de volgende evolutionaire transitie kunnen we onze aarde maar beter koesteren in plaats van uitknijpen tot de laatste druppel is verdwenen.
Een emergentie is een (verzameling van) eigenschap(pen) die schijnbaar uit het ‘niets’ opdoemt en die niet kan worden gereduceerd tot samenstellende bestanddelen. Evolutionaire emergenties lijken op priemgetallen in de zin dat ze een logisch onderdeel van een reeks uitmaken maar zich onderscheiden doordat ze niet kunnen worden ontbonden in factoren. Kenmerkend voor emergente evolutie is dat bij elke reuzensprong de oorspronkelijke elementen naast de nieuwgevormde emergentie blijven bestaan. Anders dan bijvoorbeeld het zaadje waaruit een plant groeit, blijven de kiemen van de emergentie in ons midden. Met een alfabet maak je nog geen verhaal. Voor verhalen heb je táál nodig. Zonder taal zijn verhalen onbestaanbaar. Taal is emergent.
Essentieel voor elke evolutionaire ontwikkeling is dat deze niet ophoudt bij Homo sapiens. Transhumanisme zal wellicht meer ongelijkheid met zich meebrengen, het komt voort uit een onvermijdelijk aspect van de menselijke drang naar transcendentie. Biotechnologie, cybernetica en nanotechnologie hoeven niet in strijd te zijn met de menselijke waardigheid (solidariteit, rechtvaardigheid en respect voor andersdenkenden).
Voor wie zich zorgen maakt over de toekomst, het transhumanisme zal niemand dwingen om lichaamsdelen te vervangen door superieure protheses of om zijn brein te uploaden. Pacemakers en smartphones zijn attributen waar men nu al niet meer buiten kan. De symbiose tussen mens en machine is een voortzetting van onze identiteit in wording, ons voortdurende verlangen naar meer. Het dilemma tussen angst en verlangen weerspiegelt de tweespalt tussen individu en gemeenschap en is kenmerkend voor een (volgende) emergente transitie.
Bronnen:
Jon Amsden. A History of the Earth and its Mass Extinctions. Createspace Independent Publ. Platform, 2011
Robert Axelrod. De Evolutie van Samenwerking. Contact, 1990
Hubert Van Belle & Jan van der Veken (eds.) Nieuwheid Denken. Acco, Leuven, 2008
John Bonner. First Signals. The Evolution of Multicellular Development. Princeton Univ. Press, 2000
Richard Buckminster Fuller. Synergetics: Explorations in the Geometry of Thinking. Macmillan Publ., 1982
Brett Calcott. & Kim Sterelny (eds.). The Major Transitions in Evolution Revisited. MIT Press, 2011
Hadiras Chaudhuri. The Evolution of Integral Consciousness. Wheaton, 1977; reprint Quest Books.,1989
Francis Crick. Life Itself: Its Origin and Nature. Simon & Schuster, New York, USA,1981
Charles Darwin. Over het Ontstaan van Soorten. Nieuwezijds BV, 2007
Richard Dawkins. De Zelfzuchtige Genen. Olympus Pockets, 2009
Daniel Dennet. Het Bewustzijn Verklaard. Atlas-Contact, 1999
Theodosius Dobzhansky. Evolutie en Erfelijkheid. Spectrum, 2008
Francesco Facchini. Vroegste Geschiedenis van de Mens. Veen Magazines, 2009
Brian Greene. De Ontrafeling van de Kosmos. Spectrum, 2005
Hermann Haken. Synergetics. Springer, 2004
Thomas Hartog. Het Ontstaan van de Tijd. Uitgeverij Lannoo, 2023
Stephen Hawking. Het Heelal. Bert Bakker, 1988
Bert Hölldopler & Edward Willson. The Superorganism. Norton & Comp. Inc., 2009
Fred Hoyle & Chandra Wickramasinghe. Origins of Life. Steps towards Panspermia. Kluwer Acad. Publ., 2000
Philippe Huneman & Dennis Walsh. Challenging the Modern Synthesis. Oxford Univ. Press, 2017
Stuart Kauffman. The Origins of Order. Self Organisation andSelection in Evolution. Oxford Univ. Press, 1993
Stuart Kauffman. World beyond Physics. The Emergence and Evolution of Life. Oxford Univ. Press, 2019
Jacob Klapwijk. Heeft de Evolutie een Doel? Over Schepping en Emergente Evolutie. Kok, Kampen, 2009
Elizabeth Kolbert. Het Zesde Uitsterven. Standaard Uitgeverij, 2014
Roger Lewin. Complexiteit. Contact, 1993
James Lovelock. Gaia, een nieuwe visie op de Aarde. Vbk Media, 1979
James Lovelock. De Wraak van Gaia. Bakker, 2007
Lynn Margulis. De Symbiotische Planeet. Contact, 1999
John Maynard Smith & Eörs Szathmáry. The Major Transitions in Evolution. Freeman, 1999
Carl Sagan. The Cosmic Connection.An Extraterrestial Perspective. Cambridge Univ. Press, 1973
Elliot Sober. Simplicity. Oxford University Press, UK, 1975
Gerry Webster & Brian Goodwin. Form and Transformation. Generative and Relational Principles in Biology. Cambridge Univ. Press, 1996
Ken Wilber. Sex, Ecology, Spirituality: The Spirit of Evolution. Shambala, 2001
Edward Wilson. Sociobiology. The New Synthesis. Harvard Univ. Press, 2000
Yongming Tang & Charles Joiner. Synergic Inquiry. Sage Publishing, 2006